III Международная научно-практическая конференция «Орлы Палеарктики: изучение и охрана»
Пернатые хищники и их охрана. Спецвыпуск 2. Тезисы
ГЕНЕТИЧЕСКОЕ РОДСТВО ПОПУЛЯЦИЙ ЧЁРНОГО КОРШУНА (MILVUS MIGRANS) ЮГО-ВОСТОЧНОЙ АЗИИ, ЯПОНИИ, ТАЙВАНЯ, ИНДИИ И АВСТРАЛИИ: СУЩЕСТВУЕТ ЛИ M. M. FORMOSANUS?
Андреенкова Н.Г. (Институт молекулярной и клеточной биологии СО РАН, Новосибирск, Россия)
Хун Ш.-Ю. (Институт охраны дикой природы, Колледж ветеринарной медицины, Национальный университет науки и технологий Пиндун; Тайваньская исследовательская группа по хищным птицам, Тайвань)
Линь Х.-Ш. (Институт охраны дикой природы, Колледж ветеринарной медицины, Национальный университет науки и технологий Пиндун; Тайваньская исследовательская группа по хищным птицам; Высший институт биоресурсов, Национальный университет науки и технологий Пиндун, Тайвань)
Карякин И.В. (Российская сеть изучения и охраны пернатых хищников; ООО «Сибирский экологический центр», Новосибирск, Россия)
Ивами Я. (Институт орнитологии Ямашины, Абико, Япония)
Кириллин Р.А. (Институт биологических проблем криолитозоны СО РАН, Якутск, Россия)
Литерак И. (Факультет ветеринарной гигиены и экологии, Университет ветеринарных наук Брно, Чехия)
Контакт:
Наталья Андреенкова anata@mcb.nsc.ru
Шиао-Ю Хун laughrain@gmail.com
Хуэй-Шань Линь blackkite@gmail.com
Игорь Карякин ikar_research@mail.ru
Ясуко Ивами iwami@yamashina.or.jp
Руслан Кириллин ruslan.kirillin@gmail.com
Иван Литерак LiterakI@VFU.cz
Рекомендуемая цитата: Андреенкова Н.Г., Хун Ш.-Ю., Линь Х.-Ш., Карякин И.В., Ивами Я., Кириллин Р.А., Литерак И. Генетическое родство популяций чёрного коршуна (Milvus migrans) юго-восточной Азии, Японии, Тайваня, Индии и Австралии: существует ли M. m. formosanus? — Пернатые хищники и их охрана. 2023. Спецвып. 2. С. 335-339. DOI: 10.19074/1814-8654-2023-2-335-339 URL: http://rrrcn.ru/ru/archives/35126
Ареал чёрного коршуна (Milvus migrans) захватывает обширные территории в Евразии, Африке и Австралии. Этот пернатый хищник демонстрирует уникальную экологическую пластичность, населяя самые разные биотопы и охотно используя преимущества проживания рядом с человеком.
По разным данным черный коршун имеет от пяти до семи подвидов. В Палеарктике обитают два из них: западную часть занимает европейский чёрный коршун (M. m. migrans), а восточную — черноухий коршун (M. m. lineatus). Острова Японии также населены M. m. lineatus, однако эта популяция существует достаточно изолированно от материковой и может отличаться от нее генетически. Маршруты сезонных передвижений этих птиц не выходят за пределы Японских островов, тогда как все материковые коршуны восточной Палеарктики совершают дальние миграции с севера на юг и обратно. Индию и Индокитай населяет индомалайский подвид (M. m. govinda), а в Австралии и на прилегающих к ней островах обитает (M. m. affinis). Кроме того, на территории восточной части материкового Китая, на Тайване и Хайнане в начале ХХ века был описан М. m. formosanus, напоминающий мелкую форму M. m. lineatus. С тех пор этот подвид официально присутствует во всех таксономических списках, однако он никогда прицельно не изучался. С момента описания М. m. formosanus популяции чёрного коршуна, как и многих других птиц, на этих территориях пережили резкое сокращение из-за интенсивного применения пестицидов в сельском хозяйстве. В настоящее время нет никаких данных, подтверждающих существование М. m. formosanus, как самостоятельной генетической ветви чёрного коршуна.
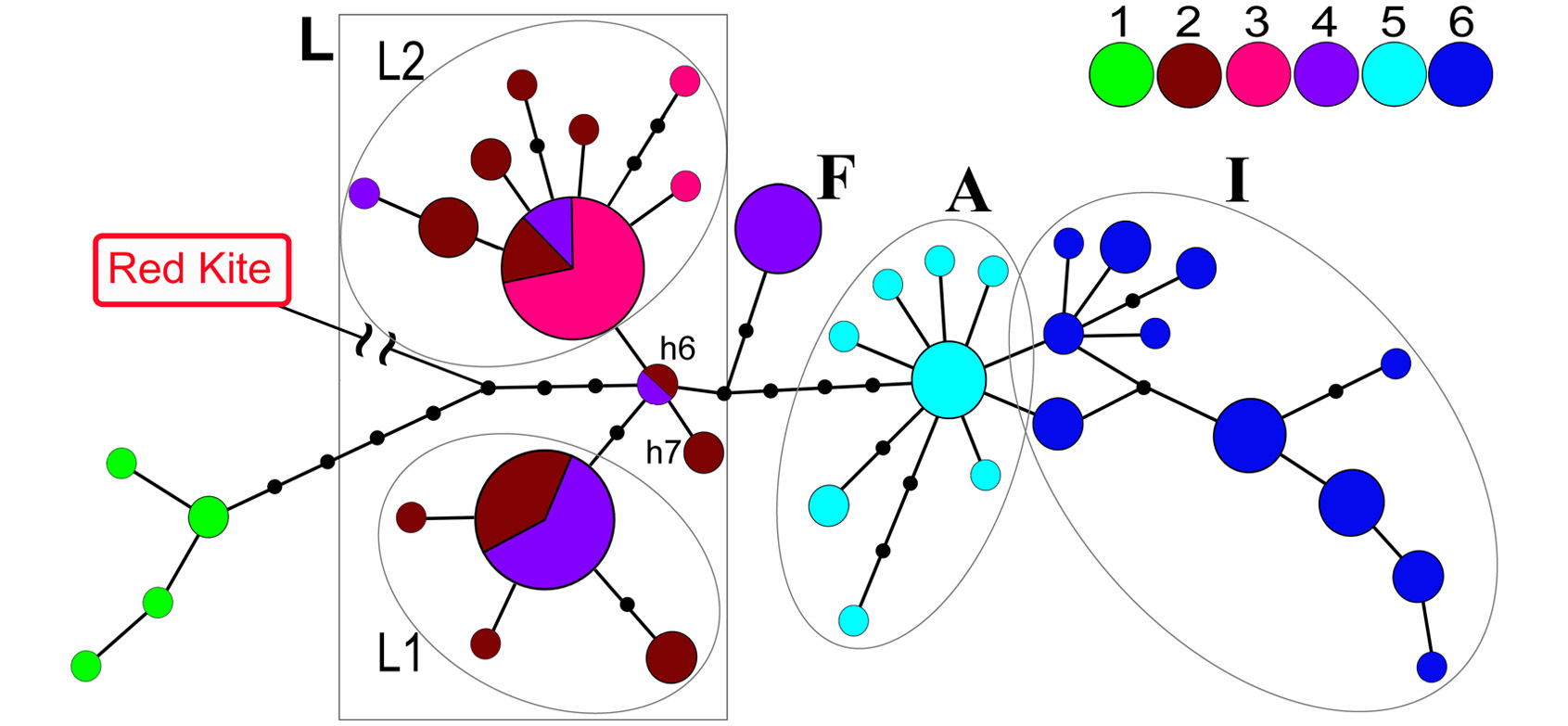
Ранее мы обнаружили, что подвиды чёрного коршуна Европы, Северной Азии и Индии отличаются генетически и имеют собственные гаплогруппы митохондриального гена цитохром В (CytB). Однако низкая вариабельность этого гена не позволила обнаружить различия между популяциями Индии и Австралии. Решив более детально исследовать восточные подвиды, мы проанализировали гены CytB и ND2 в популяциях чёрного коршуна северо-восточной Азии, Индии, Тайваня, Японии и Австралии. Оказалось, что японские коршуны несут только одну из двух основных гаплогрупп азиатских M. m. lineatus, что свидетельствует о недавней изоляции от материка. Популяции Индии и Австралии оказались очень близкими генетически, однако четко разделились по митохондриальным маркерам. По-видимому, их общий предок существовал всего около 100 тысяч лет назад, во время последнего оледенения.
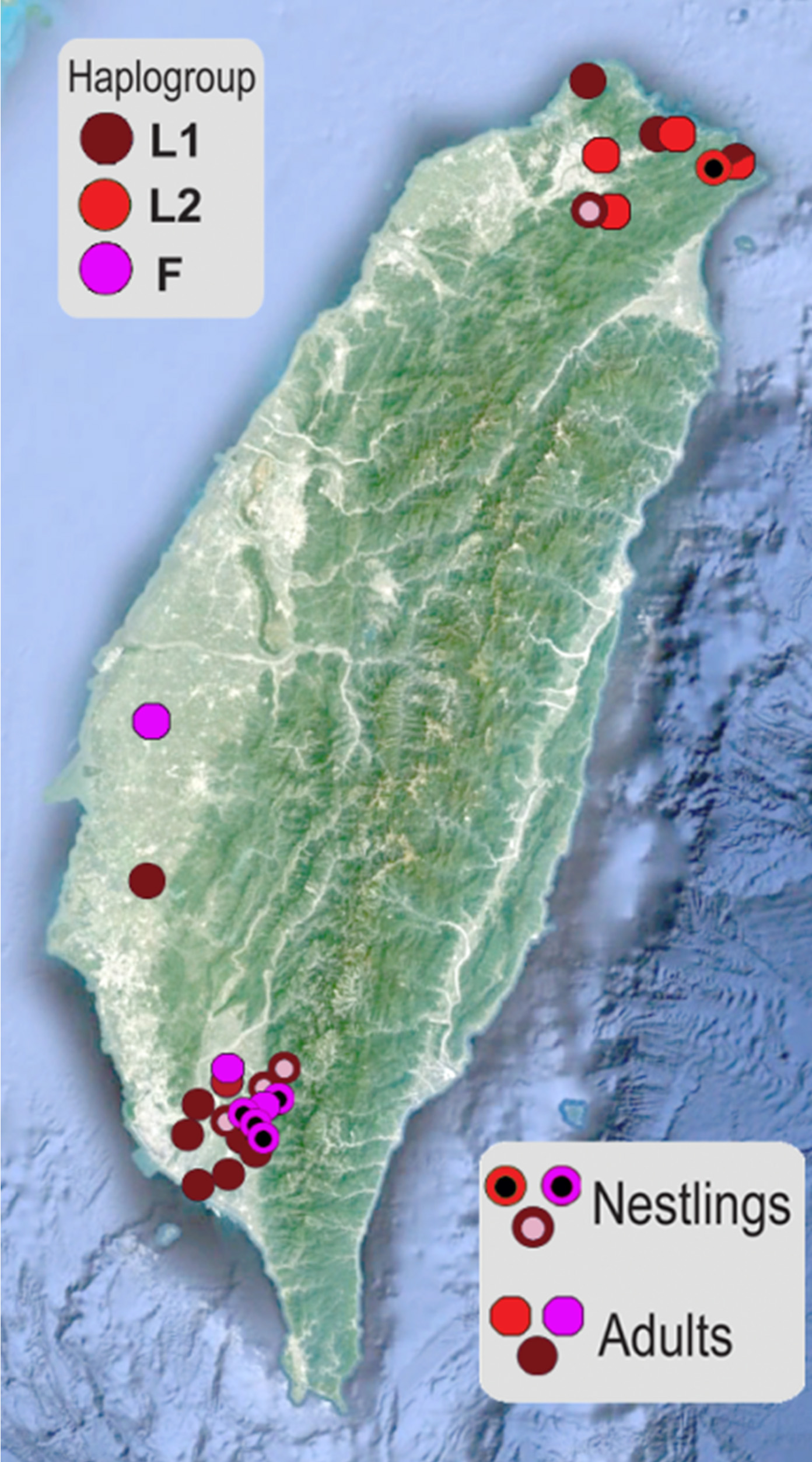
Исследуя гнездящихся коршунов Тайваня, мы обнаружили как носителей митохондриальных гаплогрупп M. m. lineatus, так и носителей новой гаплогруппы, которая до сих пор ни разу не встречалась в других популяциях. Возможно, в недавнем прошлом на острове действительно обитала самостоятельная генетическая ветвь чёрного коршуна, которая была достаточно изолированной от материка. Однако, спад численности местной популяции, а также интенсивное расселение M. m. lineatus привели к тому, что в настоящее время Тайвань населяют потомки обоих подвидов. Кроме того, резкое снижение численности коршуна в 1970-е годы, по-видимому, привело к формированию двух генетически различных популяций чёрного коршуна на юге и севере Тайваня. Существование такой изоляции при относительно небольшом расстоянии между этими популяциями вызывает удивление и требует дальнейших исследований.
Исследование выполнено за счёт гранта РНФ проект № 23-24-00185.